
Psychopathology seeks to understand the disturbances that can occur in human thought, emotion, and behavior. Among the most striking and complex of these disturbances is the loss of contact with reality. In clinical psychology and psychiatry, this phenomenon is often referred to as psychosis, a condition in which individuals experience profound alterations in perception, belief, and interpretation of the external world. When reality-testing mechanisms become impaired, the individual may develop delusions, hallucinations, or severely disorganized thinking. These experiences can fundamentally alter how a person understands themselves, other people, and the environment. Understanding why some individuals lose touch with reality requires examining biological, psychological, and social factors that interact within the human mind. Reality perception is not a simple or passive process. The human brain constantly interprets sensory information, filters it through memory and expectations, and constructs a coherent representation of the world. What individuals experience as “reality” is therefore not a direct copy of external events but a mental model created through complex cognitive processes. Under ordinary circumstances, these processes allow individuals to interpret sensory input accurately and adjust their beliefs when confronted with new evidence. However, when certain psychological or neurological mechanisms become disrupted, the brain’s model of reality can diverge significantly from the external world. One of the central mechanisms involved in maintaining contact with reality is cognitive integration. This process allows sensory information, emotional signals, and prior knowledge to be combined into a unified understanding of events. For example, when a person hears a sound, sees a visual stimulus, and recalls a related memory, the brain integrates these signals to determine what is happening. If integration functions properly, the individual forms a coherent perception of the environment. If it fails, sensory experiences may be misinterpreted or disconnected from their true sources. Hallucinations represent one of the most dramatic examples of this failure. A hallucination occurs when a person perceives something—such as a voice, image, or sensation—that does not originate from an external stimulus. In auditory hallucinations, individuals may hear voices that comment on their actions, criticize them, or carry on conversations. These voices often feel completely real to the person experiencing them, even though no physical sound exists. Research suggests that auditory hallucinations may arise when internal thoughts or verbal imagery are misidentified as external sounds. Normally, the brain distinguishes between internally generated speech and sounds coming from the outside world. This distinction involves networks responsible for monitoring one’s own mental activity. When this monitoring system becomes impaired, internal thoughts may be interpreted as external voices, creating the sensation that another entity is speaking. Delusions represent another important symptom associated with loss of reality contact. A delusion is a strongly held belief that persists despite clear evidence that it is false. Delusions can take many forms. Some individuals believe they are being followed, monitored, or persecuted. Others may believe they possess extraordinary powers, special missions, or unique connections to powerful figures. What distinguishes a delusion from ordinary belief is not simply its unusual content but the individual’s inability to reconsider it when confronted with contradictory evidence. Delusions often arise from disturbances in the brain’s systems for evaluating meaning and significance. Humans naturally search for patterns in their environment. This ability is essential for learning and survival, allowing individuals to detect relationships between events and anticipate future outcomes. However, when this pattern-detection system becomes overactive, the brain may assign excessive importance to ordinary stimuli. For example, a casual glance from a stranger might be interpreted as a sign of secret surveillance, or a random coincidence may be perceived as evidence of a hidden conspiracy. This process, sometimes called aberrant salience, occurs when neutral events are experienced as unusually meaningful or threatening. Once such interpretations form, the individual may build elaborate explanations around them, reinforcing the delusional belief. Neurochemical changes in the brain appear to play a role in this process. Dopamine, a neurotransmitter involved in motivation and reward, has been strongly linked to the development of psychotic symptoms. Excessive dopamine activity may increase the brain’s tendency to treat neutral stimuli as significant. When many unrelated events begin to feel unusually meaningful, the individual attempts to explain them, sometimes leading to the formation of delusional beliefs. While neurochemical changes can influence psychosis, biological factors alone rarely explain the full picture. Genetic vulnerability also contributes to the risk of losing contact with reality. Studies of families and twins have shown that certain psychiatric conditions involving psychosis, such as schizophrenia, tend to occur more frequently among biological relatives. This suggests that inherited differences in brain development or neurotransmitter systems may increase susceptibility. However, genetics does not determine destiny. Many individuals with genetic risk factors never develop psychosis. Environmental influences play a critical role in shaping how these vulnerabilities manifest. Stressful life events, trauma, substance use, and social isolation can all increase the likelihood that psychological symptoms will emerge. Stress is particularly important because it affects the body’s hormonal systems. When individuals experience prolonged stress, the body releases stress hormones such as cortisol. These hormones influence numerous brain regions involved in memory, emotion, and perception. Chronic elevation of stress hormones can alter neural communication and increase emotional sensitivity, potentially destabilizing the systems that regulate reality perception. Traumatic experiences may also contribute to the development of psychotic symptoms. Severe trauma—especially during childhood—can disrupt the normal development of emotional and cognitive regulation. In some cases, individuals exposed to trauma develop heightened vigilance and mistrust of their environment. This hypervigilance may lead the brain to interpret ambiguous events as threats, increasing the likelihood of paranoid thinking. Another psychological factor associated with psychosis is cognitive disorganization. In healthy cognition, thoughts follow logical connections and maintain coherent structure. During psychotic episodes, this structure may deteriorate. Speech may become fragmented or difficult to follow, and ideas may shift rapidly without clear transitions. This phenomenon reflects disruptions in the neural networks responsible for organizing language and reasoning. Cognitive disorganization can make it difficult for individuals to evaluate their own beliefs critically. When thinking processes lose coherence, the ability to compare ideas with
Why Does the Human Body Need Sleep?
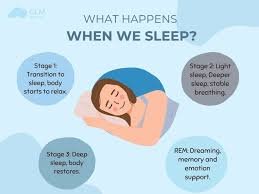
Sleep is one of the most universal biological behaviors observed across the animal kingdom. Nearly every known species—from insects to mammals—engages in some form of rest that resembles sleep. Despite this universality, sleep remains one of the most intriguing mysteries in biology. For centuries, people assumed that sleep simply allowed the body to rest after physical activity. However, modern science has revealed that sleep performs far more complex functions. During sleep, the body regulates hormones, repairs tissues, strengthens the immune system, consolidates memory, and restores energy balance. Without sleep, these processes begin to fail, affecting both physical health and cognitive functioning. Understanding why the human body requires sleep therefore requires examining the biological systems that operate during this seemingly passive state. Sleep is controlled primarily by two major regulatory systems within the body: the circadian rhythm and sleep pressure. The circadian rhythm functions as an internal biological clock that follows an approximately twenty-four-hour cycle. This clock regulates numerous physiological processes, including body temperature, hormone release, metabolism, and the timing of sleep and wakefulness. The central regulator of the circadian rhythm is a small cluster of neurons in the hypothalamus known as the suprachiasmatic nucleus. This structure receives information about light exposure through the eyes and adjusts bodily rhythms accordingly. Light plays a critical role in synchronizing the circadian clock. When light enters the retina, signals are transmitted to the suprachiasmatic nucleus, which then influences the release of hormones such as melatonin. Melatonin is produced by the pineal gland and acts as a biological signal that promotes sleep. As evening approaches and environmental light decreases, melatonin levels gradually rise, signaling to the body that it is time to prepare for rest. In the morning, exposure to light suppresses melatonin production and encourages wakefulness. The second mechanism regulating sleep is sleep pressure. Throughout the day, the body accumulates chemical signals that increase the urge to sleep. One of the most important substances involved in this process is adenosine, a molecule that builds up in the brain as energy is consumed during waking hours. As adenosine levels rise, the feeling of fatigue becomes stronger. Sleep allows the body to reduce adenosine levels, restoring alertness for the following day. Caffeine affects this system by blocking adenosine receptors in the brain. When caffeine occupies these receptors, the brain cannot detect the buildup of adenosine, temporarily reducing the sensation of fatigue. However, this effect does not eliminate sleep pressure; it merely masks it. Once the caffeine wears off, the accumulated adenosine continues to signal the need for sleep. Sleep itself occurs in several stages that repeat in cycles throughout the night. These stages are broadly divided into two categories: non-rapid eye movement sleep and rapid eye movement sleep. Non-rapid eye movement sleep includes three progressively deeper stages. In the early stage, the body begins transitioning from wakefulness to sleep. Muscle activity decreases, breathing becomes slower, and brain waves begin to change. In the deeper stages, brain activity slows significantly, and the body enters a state of profound rest. The deepest stage of sleep is particularly important for physical restoration. During this phase, the body releases growth hormone, which stimulates tissue repair, muscle development, and cellular regeneration. The immune system also becomes more active during deep sleep, producing proteins known as cytokines that help the body fight infection and inflammation. Rapid eye movement sleep represents a different physiological state. During this phase, brain activity becomes more similar to wakefulness, and vivid dreaming often occurs. Rapid eye movement sleep plays a critical role in emotional regulation and memory consolidation. Studies suggest that the brain uses this stage to process experiences from the day, integrating them into long-term memory networks. Memory formation relies on cooperation between different brain regions, particularly the hippocampus and the cerebral cortex. During waking hours, the hippocampus temporarily stores new information. While a person sleeps, especially during deep sleep stages, neural activity patterns replay these experiences, gradually transferring them to more permanent storage in the cortex. This process strengthens memories and allows individuals to retain learned information. Sleep deprivation disrupts these processes in several significant ways. When individuals do not obtain sufficient sleep, memory consolidation becomes less efficient. People may struggle to learn new information, recall recent events, or maintain concentration. Cognitive tasks requiring attention and problem-solving become more difficult as sleep loss accumulates. The immune system is also strongly influenced by sleep. Research has shown that individuals who consistently sleep less than six hours per night are more susceptible to infections. During sleep, immune cells increase their activity and produce substances that help combat viruses and bacteria. Lack of sleep reduces this activity, weakening the body’s natural defenses. Hormonal regulation is another function closely linked to sleep. Several hormones involved in appetite, metabolism, and stress depend on regular sleep patterns. For example, sleep influences the balance between two hormones that regulate hunger: leptin and ghrelin. Leptin signals feelings of fullness, while ghrelin stimulates appetite. Sleep deprivation reduces leptin levels and increases ghrelin levels, which can lead to increased hunger and overeating. This hormonal imbalance may help explain the relationship between sleep deprivation and weight gain. Individuals who regularly sleep too little often experience stronger cravings for high-calorie foods. At the same time, fatigue reduces motivation for physical activity, further contributing to metabolic imbalance. Sleep also plays a crucial role in maintaining cardiovascular health. Blood pressure normally decreases during sleep, allowing the heart and blood vessels to recover from daily activity. Chronic sleep deprivation can disrupt this process, leading to persistently elevated blood pressure and increased risk of heart disease. Another remarkable function of sleep involves the removal of metabolic waste from the brain. Recent research has identified a system known as the glymphatic system, which becomes particularly active during sleep. This system circulates cerebrospinal fluid through brain tissue, flushing away harmful waste products that accumulate during waking hours. One of these substances is beta-amyloid, a protein associated with neurodegenerative diseases when it builds up excessively. During sleep, brain cells slightly shrink in size,
Does the Brain Need Meaning to Stay Healthy?
Among the many questions explored in neuroscience, one of the most intriguing concerns the relationship between meaning and brain health. Humans do not merely react to sensory stimuli or perform mechanical behaviors; they constantly interpret their experiences through narratives of purpose, significance, and direction. People pursue careers, relationships, knowledge, and creative expression not only for survival but also for the sense that their actions matter within a broader context. This raises a profound question at the intersection of neuroscience, psychology, and philosophy: does the brain require meaning in order to function optimally, or is the human search for meaning merely a cultural construct layered upon biological processes? To explore this question, it is necessary to examine how the brain processes motivation, reward, and long-term goals. Neural systems responsible for motivation are deeply embedded in the brain’s architecture. Central among these systems is the dopaminergic reward network, which includes structures such as the ventral tegmental area, the nucleus accumbens, and parts of the prefrontal cortex. These regions work together to evaluate potential rewards and generate the motivational energy required to pursue them. Dopamine, often described as a “reward chemical,” plays a crucial role in this process. However, dopamine is not simply associated with pleasure. More accurately, it is involved in anticipation and goal-directed behavior. When the brain detects the possibility of achieving a meaningful objective—whether obtaining food, solving a complex problem, or forming a social bond—dopamine levels increase, energizing neural circuits that support effort and persistence. Meaningful goals therefore appear to influence the activity of motivational systems. When individuals perceive their actions as purposeful or valuable, the brain’s reward networks remain actively engaged. In contrast, when activities appear meaningless or disconnected from personal values, motivational circuits may become less responsive. This phenomenon is often observed in conditions characterized by loss of purpose, such as severe burnout or existential depression. From a neurological perspective, the brain evolved in environments where purposeful action was closely linked to survival. Early humans needed to hunt, gather resources, cooperate with others, and navigate complex landscapes. These tasks required sustained attention, planning, and social coordination. Neural systems developed to support these activities by reinforcing behaviors that contributed to survival and social cohesion. Meaning may therefore represent a cognitive framework that organizes motivational energy. Rather than responding only to immediate rewards, the human brain is capable of sustaining effort over long periods when actions are perceived as part of a larger narrative. This capacity relies heavily on the prefrontal cortex, the region responsible for long-term planning, abstract reasoning, and self-reflection. The prefrontal cortex allows individuals to imagine future outcomes and align present actions with those imagined possibilities. When people pursue meaningful goals—such as building a career, raising a family, or contributing to society—the brain constructs mental representations linking current behavior to future significance. These representations activate motivational circuits that sustain effort even in the absence of immediate reward. However, the brain’s need for meaning becomes particularly visible when meaning is absent. Individuals who experience prolonged states of purposelessness often report cognitive and emotional symptoms such as apathy, mental fatigue, and reduced concentration. Neuroimaging studies suggest that these states are associated with reduced activity in prefrontal motivational networks and altered dopamine signaling. Without a sense of direction, the brain may struggle to maintain engagement with complex tasks. This relationship between meaning and neural engagement also appears in educational contexts. Students who perceive their studies as relevant to personal aspirations tend to exhibit stronger attention, deeper memory formation, and greater persistence when confronted with challenging material. The hippocampus, a structure involved in learning and memory, functions more effectively when information is associated with emotional or motivational significance. In other words, meaning enhances the brain’s ability to encode and retain knowledge. The influence of meaning extends beyond cognition into emotional regulation. Emotional resilience often depends on the brain’s capacity to interpret adversity within a broader narrative framework. When individuals perceive difficulties as part of a meaningful journey—such as personal growth, moral commitment, or long-term aspiration—the emotional impact of stress may be reduced. Neural circuits involved in emotional regulation, particularly those linking the prefrontal cortex with the amygdala, appear to function more effectively under such interpretive frameworks. Conversely, when experiences appear random or meaningless, stress responses may intensify. The amygdala, responsible for detecting potential threats, becomes more reactive when situations lack clear structure or predictability. Meaning therefore provides a form of cognitive stability, allowing the brain to organize experiences into coherent patterns rather than perceiving them as chaotic or uncontrollable. Another important dimension of meaning involves social connection. Humans are inherently social organisms, and the brain contains specialized networks dedicated to understanding others’ intentions, emotions, and perspectives. These networks include regions within the medial prefrontal cortex, the temporoparietal junction, and the posterior cingulate cortex. Together, they form part of the brain’s social cognition system. Many sources of meaning emerge from social roles and relationships. Caring for family members, contributing to community life, or participating in collective goals provides a sense that one’s actions matter beyond individual survival. These experiences activate reward circuits associated with cooperation and altruism. Neurochemical processes involving oxytocin and dopamine reinforce behaviors that strengthen social bonds. Interestingly, research has shown that individuals who perceive their lives as meaningful often exhibit better physical health and longer life expectancy. While this relationship is influenced by many factors, neuroscience suggests that meaning may regulate stress physiology. People with strong purpose in life tend to display more balanced activity within the hypothalamic–pituitary–adrenal axis, the system responsible for managing stress hormones such as cortisol. Lower chronic cortisol exposure reduces the risk of inflammation, cardiovascular problems, and cognitive decline. The connection between meaning and brain health also appears in studies of aging. Older adults who maintain a strong sense of purpose often demonstrate better cognitive function and lower rates of neurodegenerative disease. One explanation is that purposeful activity encourages continuous mental engagement, which strengthens neural networks through neuroplasticity. Activities such as volunteering, mentoring, creative expression, and intellectual exploration stimulate multiple cognitive systems simultaneously. Neuroplasticity
Why Does the Brain Create Anxiety Without Danger?
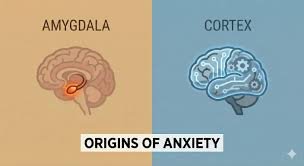
Anxiety is often understood as a rational response to threat. When a person encounters danger—a predator, an accident, or an uncertain environment—the brain activates systems that prepare the body to react quickly. Heart rate increases, attention sharpens, and muscles become ready for action. From an evolutionary perspective, this reaction is essential for survival. Yet one of the most puzzling features of human psychology is that anxiety frequently appears even when no real danger is present. Individuals may feel intense worry before a social interaction, experience fear about future events that may never occur, or sense a vague tension without any identifiable cause. This phenomenon raises a profound neurological question: why does the brain generate anxiety in the absence of immediate threat? To understand this process, it is necessary to examine the brain systems responsible for detecting and responding to danger. Central among these systems is the amygdala, a small almond-shaped structure located deep within the temporal lobes. The amygdala plays a critical role in evaluating emotional significance, particularly signals related to fear and threat. When sensory information reaches the brain, it is rapidly processed by the amygdala even before conscious awareness fully interprets the situation. This rapid processing allows the body to react to potential danger within fractions of a second. The speed of this mechanism, however, comes with an important trade-off. Because the amygdala prioritizes speed over precision, it often responds to signals that merely resemble danger rather than signals that actually represent it. From an evolutionary perspective, it is safer for the brain to mistakenly interpret a harmless stimulus as threatening than to overlook a genuine threat. A rustling sound in the bushes might be caused by wind, but if it were caused by a predator, ignoring it could be fatal. Therefore, the brain evolved to favor false alarms over missed dangers. This evolutionary bias helps explain why anxiety can arise without obvious cause. The brain constantly scans the environment for patterns associated with risk, and it does so not only through external perception but also through internal memory. Experiences from the past shape how the brain interprets present situations. If a particular environment or situation has previously been associated with embarrassment, conflict, or harm, the brain may activate anxiety even when the current circumstances are objectively safe. Memory plays a crucial role in this process. The hippocampus, a structure involved in contextual memory, works closely with the amygdala to evaluate whether a situation resembles past experiences of threat. When a similar context appears—such as a crowded room resembling a past stressful event—the hippocampus signals the amygdala, which may initiate an anxiety response. This reaction can occur even if the individual consciously knows that the current situation is not dangerous. The involvement of memory introduces an important dimension to anxiety: anticipation. Unlike many animals, humans possess a highly developed capacity for imagining future scenarios. This ability is largely mediated by networks in the prefrontal cortex, the region responsible for planning, reasoning, and simulation of possibilities. The same cognitive abilities that allow humans to prepare for future challenges also enable the brain to imagine negative outcomes in vivid detail. When the brain constructs hypothetical scenarios involving potential failure, loss, or embarrassment, the amygdala may respond as if those events were real. Physiological reactions—such as increased heart rate or muscle tension—may occur even though the imagined threat has not yet happened. In this way, anxiety can be triggered not by the external world but by the brain’s own predictions. Predictive processing has become one of the central frameworks in modern neuroscience. According to this model, the brain is not merely a passive receiver of sensory information. Instead, it actively generates predictions about what will happen next and continuously compares these predictions with incoming sensory data. When the brain detects uncertainty or mismatch between expectation and reality, it increases vigilance in order to resolve the discrepancy. Anxiety may therefore emerge when the brain perceives uncertainty about important outcomes. Situations involving social evaluation, unpredictable environments, or unclear expectations often trigger heightened predictive monitoring. The brain attempts to prepare for multiple possible outcomes simultaneously, which can produce a persistent state of tension. In such cases, anxiety functions as a motivational signal urging the individual to gather more information or regain a sense of control. Another factor contributing to anxiety without immediate danger is the brain’s sensitivity to social relationships. Human survival historically depended on cooperation within groups. Exclusion from the group could have severe consequences, including loss of protection and resources. As a result, the brain developed specialized neural circuits dedicated to monitoring social acceptance and rejection. These circuits involve regions such as the anterior cingulate cortex and the insula, both of which are associated with emotional pain. Research has shown that social rejection activates many of the same neural pathways involved in physical pain. Consequently, situations involving social evaluation—public speaking, meeting unfamiliar people, or receiving criticism—can trigger anxiety responses even though no physical threat is present. The prefrontal cortex normally helps regulate these emotional reactions by evaluating whether a perceived threat is realistic. When functioning effectively, this region can inhibit excessive amygdala activation. However, the regulatory process requires cognitive resources and energy. Under conditions of stress, fatigue, or information overload, the prefrontal cortex may become less effective at controlling emotional signals from deeper brain structures. When regulatory systems weaken, the amygdala may become more reactive. Minor uncertainties or ambiguous cues may then trigger exaggerated anxiety responses. This dynamic helps explain why individuals often experience stronger anxiety during periods of sleep deprivation, chronic stress, or emotional exhaustion. The underlying neural circuitry remains the same, but the balance between threat detection and rational evaluation shifts. Hormonal systems also contribute to the persistence of anxiety. The hypothalamic-pituitary-adrenal axis regulates the release of stress hormones such as cortisol and adrenaline. When the brain perceives potential danger, this system activates the body’s stress response, preparing muscles and cardiovascular systems for action. In short bursts, this response is adaptive and beneficial. However, when anxiety occurs repeatedly without

Can the Brain Be Tired Without Us Realizing It?
The human brain is often described as the most complex organ in the known universe. It regulates perception, emotion, reasoning, memory, and movement while silently coordinating countless physiological processes. Yet despite its extraordinary capabilities, the brain is also vulnerable to subtle forms of exhaustion that often go unnoticed. Unlike physical fatigue, which can be easily recognized through muscle soreness or a need for rest, neurological fatigue may exist quietly beneath the surface of daily functioning. Many individuals continue working, socializing, and making decisions while their cognitive systems operate under a significant hidden burden. The phenomenon of an “unnoticed tired brain” is therefore not merely metaphorical; it reflects real neurological processes that can gradually influence behavior, cognition, and emotional stability. Neurological fatigue is distinct from ordinary sleepiness. Sleepiness is typically the result of insufficient rest and can often be resolved through sleep. Neurological fatigue, however, arises when neural networks responsible for attention, regulation, and information processing become overworked over extended periods of time. In such situations, the brain does not necessarily demand immediate sleep but instead begins to function less efficiently. Decision-making slows, concentration becomes fragmented, and emotional responses may become less controlled. What makes this condition particularly complex is that the individual may not consciously notice the decline. The mind adapts to its reduced capacity and interprets it as normal functioning. Modern neuroscience has increasingly focused on the concept of cognitive load. Every moment of conscious experience requires the brain to process vast amounts of information. Sensory inputs, emotional cues, memories, and environmental signals are constantly integrated to produce coherent behavior. Under conditions of moderate load, the brain performs this task with remarkable efficiency. However, when demands exceed the system’s regulatory capacity for prolonged periods, neural efficiency begins to decline. This decline is not always dramatic. Instead, it appears gradually, manifesting as subtle lapses in attention, difficulty maintaining complex thought, or a vague sense of mental heaviness. One of the central neurological structures involved in regulating cognitive effort is the prefrontal cortex. This region, located at the front of the brain, is responsible for executive functions such as planning, impulse control, working memory, and strategic thinking. The prefrontal cortex operates as a kind of command center, coordinating information from various brain systems in order to guide behavior toward goals. When individuals engage in prolonged decision-making, intense emotional regulation, or sustained intellectual work, the metabolic demand placed on this region increases substantially. Research using functional neuroimaging has demonstrated that extended cognitive effort leads to measurable changes in prefrontal activity. Glucose consumption rises, neurotransmitter balance shifts, and neural signaling becomes less efficient. Over time, the brain begins to conserve energy by simplifying decision processes. Instead of carefully evaluating options, individuals may rely more heavily on automatic habits or emotional impulses. This transition from reflective thinking to automatic behavior is often mistaken for laziness or lack of motivation, when in reality it may reflect neurological fatigue. Another important factor contributing to unnoticed brain fatigue is the continuous stimulation characteristic of modern environments. Digital technology, social media, constant communication, and information overload expose the brain to unprecedented levels of sensory input. Unlike earlier historical periods where cognitive tasks were separated by periods of rest or routine physical labor, contemporary life often requires sustained mental engagement throughout the day. Emails, messages, notifications, and complex multitasking keep neural circuits active almost continuously. From a neurological perspective, this constant stimulation interferes with the brain’s natural rhythms of activation and recovery. Neural systems function most effectively when periods of effort alternate with moments of quiet processing. During rest, the brain activates what neuroscientists call the “default mode network.” This network is involved in internal reflection, memory consolidation, and emotional integration. Without sufficient time in this restorative mode, neural circuits remain in a prolonged state of task-oriented activity, increasing the risk of hidden fatigue. One of the most fascinating aspects of neurological fatigue is its relationship with emotional processing. The brain structures responsible for emotion regulation, including the amygdala and anterior cingulate cortex, are closely connected to executive systems in the prefrontal cortex. When cognitive fatigue develops, the regulatory influence of the prefrontal cortex over emotional centers becomes weaker. As a result, emotional reactions may become slightly more intense, less filtered, or more unpredictable. This does not necessarily mean that individuals experience dramatic emotional outbursts. More often, the changes are subtle. A person may become slightly more irritable, less patient in conversation, or more sensitive to minor frustrations. Because these changes emerge gradually, they are rarely attributed to neurological fatigue. Instead, they are interpreted as personality fluctuations or reactions to external circumstances. Memory performance also changes under conditions of hidden brain fatigue. The hippocampus, a structure crucial for forming new memories, relies on coordinated interaction with attention networks in the prefrontal cortex. When cognitive fatigue weakens attentional control, the brain becomes less effective at encoding new information. People may notice that they reread the same paragraph multiple times, forget small details, or struggle to recall names or recent events. Again, these symptoms are usually mild and therefore easily dismissed. Long-term exposure to unnoticed neurological fatigue may influence decision-making patterns in more profound ways. Decision neuroscience has shown that when cognitive resources are depleted, individuals tend to prefer simpler choices and immediate rewards rather than complex long-term strategies. This phenomenon has been observed in numerous behavioral experiments. Participants performing demanding cognitive tasks often shift toward more impulsive decision patterns afterward. The brain, in effect, attempts to conserve energy by avoiding cognitively expensive reasoning. Interestingly, neurological fatigue does not always feel unpleasant. In some cases, individuals describe a strange sense of emotional flatness or detachment rather than exhaustion. The brain reduces its responsiveness in order to maintain stability. This reduction may produce a temporary feeling of calm or numbness, masking the underlying cognitive depletion. Because the sensation is not strongly negative, people may continue functioning without recognizing the need for recovery. Sleep plays a crucial but complex role in the recovery of neural energy. During deep sleep, the brain undergoes

Is Loneliness a Neurological Pain?
Loneliness is commonly described as an emotional state, a subjective feeling that arises when a person’s social connections do not meet their psychological needs. However, over the past several decades, research in neuroscience has increasingly suggested that loneliness is not merely an abstract emotional experience. Instead, it may function as a biologically embedded signal deeply rooted in the human nervous system. Some neuroscientists even argue that loneliness resembles a form of neurological pain, sharing mechanisms with the neural circuits that process physical suffering. This perspective reframes loneliness not simply as sadness or isolation, but as a neurobiological warning system designed to protect human survival. Human beings evolved as profoundly social organisms. Throughout evolutionary history, survival depended heavily on cooperation, shared resources, and collective protection. Being excluded from a group or separated from social bonds significantly reduced an individual’s chances of survival. Consequently, natural selection likely favored neurological mechanisms that would strongly motivate individuals to maintain social connections. Just as physical pain warns the body of injury or danger, loneliness may operate as a psychological alarm that alerts the brain to social disconnection. Neuroscientific research has identified several brain regions associated with the experience of loneliness. Among these regions, the anterior cingulate cortex plays a particularly important role. This area of the brain is involved in processing both emotional distress and physical pain. Functional neuroimaging studies have demonstrated that when individuals experience social rejection or feelings of exclusion, activity increases in the same neural circuits that are activated during physical pain. In other words, the brain appears to process social pain and physical pain through overlapping neurological systems. This overlap may explain why loneliness can feel physically uncomfortable. Individuals experiencing prolonged loneliness often describe sensations such as chest tightness, bodily heaviness, or fatigue. These physical experiences are not merely metaphors. Instead, they reflect the activation of stress-related neural and hormonal systems within the body. The brain interprets social isolation as a potential threat, triggering physiological responses designed to restore safety and connection. One of the primary biological systems involved in this process is the hypothalamic-pituitary-adrenal axis, often abbreviated as the HPA axis. When the brain perceives stress or threat, the HPA axis activates the release of cortisol, a hormone that prepares the body to respond to danger. In short-term situations, this response is adaptive and beneficial. However, chronic loneliness can lead to prolonged activation of the stress system, resulting in persistently elevated cortisol levels. Elevated cortisol has widespread effects on the body and brain. It can disrupt sleep patterns, weaken immune functioning, and impair cognitive performance. Long-term exposure to high cortisol levels has also been associated with structural changes in the hippocampus, a brain region crucial for memory formation and emotional regulation. Thus, loneliness does not only affect emotional wellbeing; it can gradually reshape neural architecture and physiological health. Another key neurochemical system involved in loneliness is the dopamine reward system. Social interaction typically activates dopamine pathways in the brain, producing feelings of pleasure, motivation, and reinforcement. Positive social contact essentially acts as a natural reward that encourages individuals to maintain relationships and cooperative behaviors. When social contact is absent, the reward system receives less stimulation, leading to decreased motivation and emotional flattening. This reduction in dopamine activity may partly explain why chronic loneliness often leads to symptoms resembling depression. Individuals who feel socially disconnected frequently report diminished pleasure in activities, low motivation, and emotional numbness. These symptoms reflect the reduced activation of neural reward circuits that normally respond to social engagement. In addition to dopamine, the neuropeptide oxytocin plays a crucial role in regulating social bonding. Oxytocin is often referred to as the “bonding hormone” because it is released during positive social interactions such as physical touch, eye contact, and emotional intimacy. Oxytocin promotes trust, reduces fear responses, and strengthens interpersonal attachment. When individuals lack meaningful social contact, oxytocin activity decreases, potentially intensifying feelings of isolation and mistrust. This reduction in oxytocin may also contribute to a paradoxical pattern observed in chronic loneliness. While lonely individuals desire social connection, they often become more socially withdrawn or hypervigilant in social situations. Neuroscientists propose that prolonged isolation sensitizes the brain’s threat-detection systems, particularly within the amygdala. As a result, lonely individuals may become more sensitive to potential rejection or negative social cues. This heightened sensitivity can create a self-reinforcing cycle. Because lonely individuals anticipate rejection, they may interpret ambiguous social signals as negative or threatening. These interpretations can lead to avoidance behaviors, reducing opportunities for positive interaction. Over time, the absence of rewarding social experiences further strengthens feelings of isolation and reinforces the neural patterns associated with loneliness. Another important dimension of loneliness involves the brain’s default mode network, a system of interconnected regions that becomes active when the mind is not focused on external tasks. The default mode network is heavily involved in self-reflection, autobiographical memory, and social cognition. Studies have shown that lonely individuals often exhibit altered activity within this network, particularly in regions responsible for thinking about others’ intentions and social relationships. This altered activity may increase rumination and self-focused thought patterns. Rather than facilitating adaptive social reflection, the default mode network may become dominated by repetitive concerns about rejection, inadequacy, or social failure. These cognitive loops can intensify emotional distress and make it more difficult for individuals to engage in new social experiences. The neurological effects of loneliness also extend into the immune system. Research in psychoneuroimmunology has demonstrated that chronic loneliness is associated with increased inflammatory responses in the body. Specifically, lonely individuals often exhibit elevated levels of pro-inflammatory cytokines, molecules that play a role in the body’s immune defense. Inflammation is beneficial when responding to injury or infection, but chronic inflammation can contribute to numerous health problems, including cardiovascular disease, metabolic disorders, and neurodegenerative conditions. Some researchers believe that the immune system evolved to interpret social isolation as a potential signal of environmental danger. Historically, individuals separated from their group may have faced greater risk of injury or infection, prompting the immune system to adopt

Can the Brain Change Itself Through Thought?
Can deliberate thought reshape the physical structure of the brain, or are neural pathways fixed once development is complete? For much of scientific history, the prevailing assumption held that the adult brain was largely static. According to this earlier view, neurons that were damaged could not regenerate, and neural circuits established during childhood remained fundamentally unchanged throughout life. However, advances in neuroscience during the late twentieth and early twenty-first centuries have radically transformed this perspective. The concept of neuroplasticity—the brain’s capacity to reorganize its structure and function in response to experience—has become one of the most significant discoveries in modern cognitive science. Within this framework emerges a deeper question: can intentional mental activity, such as focused attention, reflection, or meditation, actively reshape the brain itself? The human brain is composed of approximately eighty-six billion neurons interconnected through trillions of synaptic connections. These connections are not static. They strengthen, weaken, and reorganize depending on patterns of activity. When particular neural pathways are repeatedly activated, the synapses connecting those neurons become more efficient through processes such as long-term potentiation. Conversely, pathways that are rarely used may weaken or disappear. This dynamic process reflects the brain’s fundamental principle: structure follows activity. Experience plays a central role in shaping neural architecture. Learning a new language, practicing a musical instrument, or mastering a complex skill results in measurable structural changes within relevant brain regions. For instance, musicians often exhibit enlarged cortical representations associated with fine motor control and auditory processing. Similarly, individuals who engage in intensive spatial navigation, such as taxi drivers navigating complex urban environments, show increased hippocampal volume associated with spatial memory. These findings demonstrate that repeated behavioral experience modifies neural organization. Yet behavioral experience is not limited to physical actions. Thought itself represents a form of neural activity. When individuals focus attention, rehearse memories, or imagine future scenarios, specific neural circuits become active. Over time, repeated patterns of mental activity may strengthen the underlying neural pathways. In this sense, the boundary between thought and behavior becomes blurred; both involve patterns of neural firing capable of reshaping the brain. Meditation research provides some of the most compelling evidence supporting the capacity of thought to alter neural structure. Long-term practitioners of mindfulness and contemplative meditation display measurable differences in brain regions associated with attention regulation, emotional processing, and self-awareness. Neuroimaging studies have shown increased cortical thickness in areas of the prefrontal cortex and anterior cingulate cortex among experienced meditators. These regions are associated with executive control and emotional regulation. The repeated practice of directing attention inward appears to strengthen networks responsible for monitoring and regulating mental states. Furthermore, meditation has been associated with changes in the amygdala, a structure involved in fear and stress responses. Individuals who engage in sustained contemplative practice often show reduced amygdala activation in response to emotional stimuli, suggesting enhanced emotional regulation. These findings imply that intentional cognitive practices can reshape neural circuits related to emotional reactivity. Rather than being passive recipients of neural processes, individuals may participate in shaping those processes through disciplined mental training. Cognitive behavioral therapy (CBT) offers another example of thought influencing neural architecture. CBT encourages individuals to identify and modify maladaptive patterns of thinking that contribute to anxiety, depression, or other psychological disorders. Neuroimaging studies indicate that successful therapy correlates with changes in brain activity within circuits involving the prefrontal cortex and limbic system. As patients learn to reinterpret negative experiences and challenge automatic thoughts, neural patterns associated with emotional regulation appear to shift. These findings suggest that cognitive restructuring—the deliberate modification of thought patterns—can produce measurable biological effects. The mechanism underlying these changes involves synaptic plasticity. When particular thoughts are repeated, the neurons participating in those thought patterns fire together repeatedly. According to Hebbian theory, neurons that fire together wire together. Over time, these repeated activations strengthen the synaptic connections linking them. Thus, habitual thought patterns—whether constructive or destructive—can gradually sculpt neural architecture. This principle carries important implications for mental health. Persistent negative rumination may reinforce neural circuits associated with anxiety or depressive thinking. Conversely, practices that cultivate gratitude, compassion, or positive reinterpretation may strengthen alternative networks promoting resilience and emotional balance. While such changes do not occur instantly, consistent mental practice can gradually alter the probability of certain patterns of thought arising spontaneously. Attention plays a particularly crucial role in this process. The brain allocates resources preferentially to stimuli receiving focused attention. When attention is directed toward specific experiences, the neural representations associated with those experiences become more strongly encoded. In effect, attention acts as a gatekeeper determining which neural pathways are reinforced. Practices that cultivate sustained attention—such as mindfulness meditation—may therefore enhance the brain’s capacity for self-directed plasticity. The role of imagination further illustrates the power of thought. Studies have shown that mentally rehearsing a physical action can activate neural circuits similar to those involved in performing the action itself. Athletes often use visualization techniques to improve performance, mentally practicing movements before executing them physically. Neuroimaging reveals that such visualization engages motor planning regions of the brain, reinforcing neural pathways associated with the imagined activity. Over time, this mental rehearsal can improve real-world performance. However, the capacity of thought to reshape the brain is not unlimited. Biological constraints still apply. Genetic factors influence neural development, and certain structural changes require extended periods of consistent practice. Moreover, the brain’s plasticity varies across the lifespan. During early childhood, neural circuits are especially malleable. As individuals age, plasticity decreases, though it never disappears entirely. Even in later adulthood, learning and mental training can induce measurable neural changes. Another important limitation concerns the difference between short-term neural activation and long-term structural change. A single episode of focused thought may temporarily activate certain neural circuits, but lasting structural modification typically requires repeated practice over extended periods. Neuroplasticity is cumulative rather than instantaneous. The brain adapts gradually as patterns of activity become habitual. Emerging research also explores how emotional states interact with neuroplasticity. Positive emotional engagement appears to facilitate learning and neural adaptation, whereas chronic stress may

Is the Self an Illusion Constructed by the Brain?
Is the self a stable, enduring entity that persists through time, or is it an illusion generated by neural processes to create coherence out of complexity? This question penetrates to the core of philosophy, neuroscience, psychology, and even contemplative traditions. The sense of being a unified “I” — a subject of experience who thinks, decides, remembers, and acts — feels immediate and undeniable. Yet contemporary research increasingly suggests that this sense of unity may be a construction rather than a metaphysical fact. If the self is a construct, what does that mean for identity, responsibility, continuity, and meaning? At the level of lived experience, the self appears singular and continuous. One remembers childhood, anticipates the future, and feels ownership over bodily movements and thoughts. This continuity fosters the belief that there exists an inner subject — a core identity — persisting unchanged despite external transformations. Classical philosophy often reinforced this intuition. From Descartes’ cogito to Kant’s transcendental unity of apperception, the self was treated as foundational: the necessary condition for coherent experience. However, empirical investigation complicates this picture. Neuroscience reveals no single location in the brain corresponding to a centralized “self module.” Instead, self-related processes are distributed across multiple networks, including regions involved in memory, interoception, emotional evaluation, and social cognition. These networks coordinate to produce the experience of unity, yet none alone embodies a permanent essence. The absence of a neural homunculus — a central observer inside the brain — challenges the notion of a fixed internal agent. The brain integrates sensory input, bodily signals, and autobiographical memory into a coherent narrative. This integration is dynamic and context-dependent. Damage to particular neural systems can fragment aspects of identity. For example, disruptions in memory can impair continuity, while disturbances in interoceptive processing can alter bodily ownership. Conditions such as depersonalization disorder illustrate how fragile the sense of self can be. Individuals may report feeling detached from their own experiences, as if observing life from a distance. Such phenomena suggest that the self is not a metaphysical constant but a neurocognitive achievement. Split-brain research further illuminates the constructed nature of selfhood. When the corpus callosum connecting the brain’s hemispheres is severed, each hemisphere can process information independently. In certain experimental contexts, patients appear to exhibit dual streams of awareness. Yet rather than experiencing themselves as two separate persons, many individuals maintain a unified sense of identity. The brain’s interpretive mechanisms generate coherence even when underlying processing is divided. This indicates that unity may be imposed retrospectively through narrative integration. The narrative theory of identity posits that the self is fundamentally a story the brain tells about itself. Autobiographical memory organizes experiences into a temporally extended account, selecting and interpreting events to support coherence. The self, in this view, is not discovered but composed. Memory is reconstructive, not archival; each act of recollection reshapes the narrative. As beliefs and values evolve, past experiences are reinterpreted. The self becomes an ongoing project rather than a static core. Psychological research supports this narrative perspective. Individuals routinely misremember events in ways that reinforce their current self-concepts. Traits perceived as central to identity influence which memories are retained and how they are framed. The brain favors coherence over contradiction. Inconsistent experiences are often rationalized or forgotten. This selective reconstruction stabilizes identity while simultaneously demonstrating its malleability. The predictive processing framework offers another angle. According to this theory, the brain functions as a hierarchical prediction machine, constantly generating models to anticipate sensory input. The self may represent one such model — a predictive construct that organizes perception and action. By attributing experiences to a unified agent, the brain simplifies complex processes. The sense of ownership over thoughts and actions could emerge from successful predictive integration rather than from an independent entity. Embodiment plays a crucial role in this construction. Interoceptive signals from the body — heartbeat, respiration, visceral sensations — contribute to the feeling of being a localized subject. Experiments such as the rubber hand illusion demonstrate how easily bodily ownership can be manipulated. When visual and tactile cues are synchronized, individuals can experience a fake hand as their own. This malleability indicates that bodily self-representation depends on multisensory integration rather than on immutable identity. Social cognition also shapes the self. Humans develop identity within relational contexts. Language, cultural norms, and interpersonal feedback influence self-concept. The brain encodes representations of how others perceive us, integrating these perspectives into internal narratives. The self is thus partially constituted by social interaction. Without relational input, identity formation would be radically altered. This interdependence challenges the idea of a self-contained, autonomous core. Philosophically, the notion of self as illusion echoes certain strands of Buddhist thought, which argue that what we call the self is a composite of transient processes — sensations, perceptions, thoughts — lacking inherent substance. Contemporary neuroscience does not adopt metaphysical claims about non-self, yet it converges on a similar functional insight: unity arises from dynamic aggregation rather than from an enduring essence. However, declaring the self an “illusion” risks misunderstanding. Illusions are typically false perceptions masking underlying reality. If the self is illusory, what is the reality beneath it? Some argue that the term “construction” is more accurate. The self may be a real process, though not a static object. Just as a whirlpool exists as a dynamic pattern in water rather than as a separate substance, the self might exist as a pattern of neural activity persisting through change. The ethical implications of this perspective are profound. Responsibility presupposes agency, and agency presupposes a self. If the self is merely a construct, does accountability dissolve? Compatibilist interpretations suggest otherwise. Even if identity is constructed, the processes generating it remain organized and coherent enough to support moral evaluation. The absence of metaphysical substance does not negate functional continuity. Memory and anticipation further sustain the sense of persistence. Episodic memory connects past experiences to present identity, while prospection projects the self into imagined futures. Neuroimaging studies reveal overlapping neural circuits for remembering and imagining, indicating that temporal extension
Does Free Will Survive Neuroscience?
Does free will endure under the scrutiny of contemporary neuroscience, or does empirical investigation dissolve the very notion of autonomous agency? Few questions generate as much tension between scientific discovery and philosophical tradition as this one. For centuries, free will has occupied a central place in moral theory, law, theology, and personal identity. It underwrites responsibility, praise, blame, and the sense that one could have acted otherwise. Yet advances in neuroscience increasingly reveal the neural mechanisms underlying decision-making, suggesting that choices arise from processes operating prior to conscious awareness. If brain activity determines behavior before conscious intention emerges, can freedom remain anything more than an illusion? The classical conception of free will presupposes that agents possess the capacity to initiate actions independent of deterministic chains of causation. In this libertarian framework, freedom entails genuine alternative possibilities. One is not merely the endpoint of prior causes but an originator of action. This view aligns with intuitive experience: when deliberating between options, individuals feel that they could choose differently. However, neuroscience challenges the reliability of this introspective certainty. Experimental studies have demonstrated that neural activity predictive of a motor decision can be detected milliseconds before participants report conscious intention to act. Such findings suggest that the brain prepares actions prior to the subjective awareness of deciding. If conscious intention follows rather than precedes neural initiation, then conscious will may not be the causal driver it appears to be. Instead, awareness might function as a post hoc narrative, integrating actions already set in motion by unconscious processes. Deterministic interpretations of these findings argue that free will is incompatible with neurobiological causation. According to this view, every decision results from prior neural states shaped by genetics, environment, and developmental history. The brain operates according to physical laws; therefore, human behavior is ultimately determined. The feeling of freedom may be a byproduct of complex information processing rather than evidence of metaphysical independence. Yet the inference from neural precursors to the negation of free will is not straightforward. First, the temporal gap between neural preparation and conscious awareness does not necessarily imply that conscious processes are causally irrelevant. Conscious deliberation may occur earlier in the cognitive sequence, influencing neural configurations long before specific motor preparations are detectable. Moreover, milliseconds of precedence do not conclusively establish determinism; they merely reveal that conscious intention is embedded within broader neural dynamics. Compatibilist philosophers argue that free will does not require exemption from causation. Instead, freedom consists in acting according to one’s reasons, values, and deliberative capacities without external coercion. On this account, the fact that decisions arise from neural processes does not undermine freedom, because those processes constitute the agent. The brain is not an alien force imposing actions upon the self; it is the biological substrate of the self. To say that neural mechanisms determine behavior is not to say that the agent is absent. Neuroscience also reveals the complexity and plasticity of decision-making systems. The prefrontal cortex integrates information about goals, social norms, and long-term consequences. Damage to these regions can impair impulse control and moral reasoning, indicating that free agency depends upon intact neural circuits. Rather than eliminating free will, such findings underscore its embodied basis. Agency emerges from distributed neural networks coordinating perception, memory, and valuation. Predictive processing theories further complicate simplistic determinism. The brain continuously generates predictions about sensory input and updates them through feedback. Decision-making involves evaluating competing predictions and minimizing error signals. This dynamic process includes both automatic and reflective components. Conscious deliberation may serve as a higher-level regulatory mechanism, modulating lower-level impulses. Even if underlying computations follow physical laws, the hierarchical organization of these processes can sustain meaningful forms of self-control. Quantum indeterminacy is sometimes invoked to rescue libertarian freedom. If neural processes incorporate indeterminate events, perhaps choices are not strictly determined. However, randomness does not equate to freedom. Actions resulting from chance fluctuations would not be more autonomous than those determined by prior causes. The challenge is not merely to avoid determinism but to account for rational control and responsibility. The legal and ethical implications of neuroscience are profound. If behavior is entirely determined by neural states beyond conscious control, retributive punishment loses justification. Responsibility might shift toward prevention and rehabilitation. Yet most legal systems operate under a compatibilist assumption: individuals are responsible when they possess the capacity for rational deliberation and are not subject to coercion or severe impairment. Neuroscience refines understanding of these capacities without necessarily abolishing them. Some neuroscientists caution against overinterpreting experimental data. Laboratory tasks involving simple motor decisions differ substantially from complex moral choices. Pressing a button at an arbitrary moment may rely heavily on automatic neural processes, whereas deliberating about career changes or ethical dilemmas engages extended reflective reasoning. Generalizing from simplified paradigms to all forms of agency risks reductionism. Furthermore, conscious intention may not function as a singular event but as an ongoing process distributed over time. Rather than a discrete moment of willing, agency may consist in sustained patterns of self-regulation. The brain’s readiness potentials could reflect preliminary fluctuations within a larger decision architecture shaped by prior commitments and goals. Conscious awareness might represent the culmination of earlier evaluative activity rather than an afterthought. The phenomenology of freedom also warrants examination. The experience of choosing involves a sense of ownership and authorship. Even if neural events precede awareness, the integration of these events into coherent self-representation may constitute genuine agency. The self is not an entity separate from neural processes but a pattern instantiated by them. To dismiss agency because it arises from neural activity would be analogous to dismissing life because it arises from biochemistry. Philosophical skepticism persists, however. If every choice can be traced to prior causes extending beyond individual control—genes inherited, environments encountered—then ultimate responsibility appears elusive. One did not choose one’s genetic endowment or early upbringing, yet these factors profoundly shape decision tendencies. The regress of causation challenges the notion of self-origination. Compatibilism responds by redefining responsibility in relational terms. Moral evaluation concerns whether actions express the
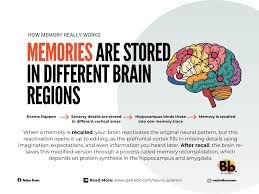
Is Memory a Reconstruction Rather Than a Record?
Is memory a faithful archive of lived experience, or is it an active reconstruction shaped by present needs, beliefs, and emotional states? The intuitive metaphor of memory as a recording device—storing events as they occurred and retrieving them intact—has long dominated common understanding. Yet contemporary cognitive science, neuroscience, and philosophy increasingly challenge this archival model. Evidence suggests that memory is not a static record but a dynamic, reconstructive process. To remember is not merely to retrieve the past but to recreate it within the interpretive frameworks of the present. The archival model of memory implies stability, durability, and fidelity. In this view, experiences are encoded, stored, and later accessed without substantial alteration. Such a model aligns with everyday language: we “store” memories, “retrieve” them, and “lose” them. However, empirical findings undermine this metaphor. Psychological research has repeatedly demonstrated that recollections are susceptible to distortion through suggestion, emotional reinterpretation, and contextual reframing. Individuals can develop vivid memories of events that never occurred, particularly when exposed to misleading information. These findings indicate that memory is malleable rather than fixed. Neuroscientific research reinforces this perspective. Memory formation involves complex interactions among the hippocampus, prefrontal cortex, and distributed cortical networks. Encoding does not produce a literal imprint of experience but rather a pattern of neural activation that is reassembled during recall. Each act of remembering reactivates and potentially modifies the memory trace, a process known as reconsolidation. During reconsolidation, memories become temporarily labile and subject to alteration before being stored again. This cyclical updating mechanism suggests that memory is continuously rewritten rather than preserved in original form. The reconstructive nature of memory also reflects its adaptive function. From an evolutionary standpoint, the purpose of memory may not be precise preservation but flexible guidance for future behavior. Accurate recall of every detail would be computationally inefficient and potentially maladaptive. Instead, memory abstracts patterns, extracts meaning, and integrates new information. Such abstraction enables generalization and problem-solving but inevitably sacrifices exactitude. Memory prioritizes coherence and relevance over literal accuracy. Emotion exerts a powerful influence on reconstruction. Emotional arousal enhances the consolidation of certain aspects of experience while narrowing attention to central features. Peripheral details may fade or transform. Over time, the emotional tone associated with a memory can shift, altering interpretation. For instance, an event initially experienced as humiliating may later be reframed as formative or humorous. The remembered event thus reflects ongoing narrative integration rather than immutable fact. Social interaction further shapes memory reconstruction. Recollections are often shared, negotiated, and co-constructed within communities. Collective memory emerges through repeated retelling, with each iteration subtly modifying content. Social conformity pressures can influence individual recall, leading people to align their memories with group consensus. Memory, therefore, is not solely an internal cognitive process but a socially embedded phenomenon. Language plays a crucial mediating role. Verbal encoding transforms perceptual experience into symbolic representation. The vocabulary and conceptual categories available at the time of recall influence reconstruction. Descriptive framing can alter perceived details; for example, wording that implies intensity or causation can change how an event is remembered. This linguistic modulation demonstrates that memory is filtered through interpretive schemas. The phenomenon of autobiographical memory illustrates the narrative dimension of reconstruction. Individuals construct life stories that provide continuity and identity. Memories are selected and organized to support coherent self-concepts. Inconsistencies are often resolved through reinterpretation rather than retention of contradiction. The self is not merely shaped by memory; memory is shaped by the evolving self. As beliefs and values change, recollections are subtly reconfigured to maintain narrative alignment. Neuropsychological conditions provide additional insight. Patients with amnesia often retain procedural memory while losing episodic recall, indicating that different memory systems operate under distinct principles. Conversely, individuals with hyperthymesia—exceptionally detailed autobiographical recall—still exhibit distortions upon close examination. Even seemingly exact memory does not escape reconstructive influence. These cases suggest that reconstruction is intrinsic to memory architecture rather than a flaw in particular individuals. Philosophical analysis deepens the inquiry. If memory is reconstructive, what becomes of epistemic reliability? Knowledge of the past depends upon trustworthy recollection. A purely reconstructive account might appear to undermine confidence in memory as a source of evidence. However, reconstruction does not entail radical unreliability. Cognitive systems evolved to balance flexibility with accuracy. While specific details may shift, core structures often remain stable. Memory is probabilistic rather than arbitrary. The relationship between memory and imagination further complicates the distinction. Neuroimaging studies reveal overlapping neural substrates for remembering the past and imagining the future. Both involve constructive simulation. This overlap suggests that memory may function as a database of elements recombined to anticipate possibilities. In this framework, recollection and projection share cognitive mechanisms. Memory becomes a form of constrained imagination grounded in prior experience. Trauma research highlights both the strengths and vulnerabilities of reconstructive memory. Traumatic memories can be intrusive and vividly sensory, yet they may also be fragmented or dissociated. Therapeutic interventions often involve narrative reconstruction, enabling individuals to recontextualize and integrate traumatic experiences. The possibility of therapeutic modification underscores the plasticity of memory but also raises ethical questions regarding suggestibility and false recollections. Technological metaphors frequently mislead understanding. Comparing memory to digital storage implies discrete files retrievable without alteration. Biological memory operates differently. Neural networks encode information in distributed patterns sensitive to context. Recall depends on cues that activate partial representations, which are then completed through inference. This inferential completion is inherently reconstructive. Rather than retrieving a complete record, the brain reconstructs a plausible scenario consistent with available traces. Developmental psychology demonstrates that reconstructive processes emerge early. Children’s memories are particularly susceptible to suggestion, yet adults remain vulnerable as well. Over time, repeated storytelling can solidify altered versions of events, making them subjectively indistinguishable from original perceptions. Confidence in memory does not reliably correlate with accuracy, revealing a gap between subjective conviction and objective fact. The reconstructive model also aligns with predictive processing theories of cognition. According to these theories, the brain continuously generates predictions about sensory input and updates them through error correction. Memory contributes to predictive models