Memory is often treated as a stable archive within the nervous system, a place where experiences are stored and later retrieved when needed. This metaphor suggests a kind of neurological library in which events, knowledge, and emotions are deposited in organized form. Yet modern neuroscience challenges this simplified view. The brain does not contain a single location where memories reside as intact objects. Instead, memory emerges from complex patterns of neural activity distributed across multiple systems. The question therefore arises in a deeper sense: where is memory actually located?
The difficulty of answering this question begins with the nature of neural representation. When an event occurs, sensory systems transform external stimuli into electrical signals. Visual structures process patterns of light, auditory regions decode sound frequencies, and somatosensory networks register touch and bodily position. Each modality contributes fragments of information about the same event.
These fragments do not converge into a single unified storage site. Instead, they remain partially distributed across the cortical regions responsible for their original processing. The color of an object, the sound associated with it, the emotional tone of the situation, and the spatial environment in which it occurred are represented by overlapping neural circuits.
Memory therefore depends on the capacity of the brain to reactivate these distributed patterns in coordinated ways. Retrieval does not involve opening a mental container where a complete record has been preserved. Rather, it requires reconstructing an approximation of the original neural configuration.
This reconstruction process explains why memory is often imperfect. Each act of remembering partially rebuilds the past using fragments stored in different networks. During this process, new interpretations and present emotional states may influence how those fragments are combined.
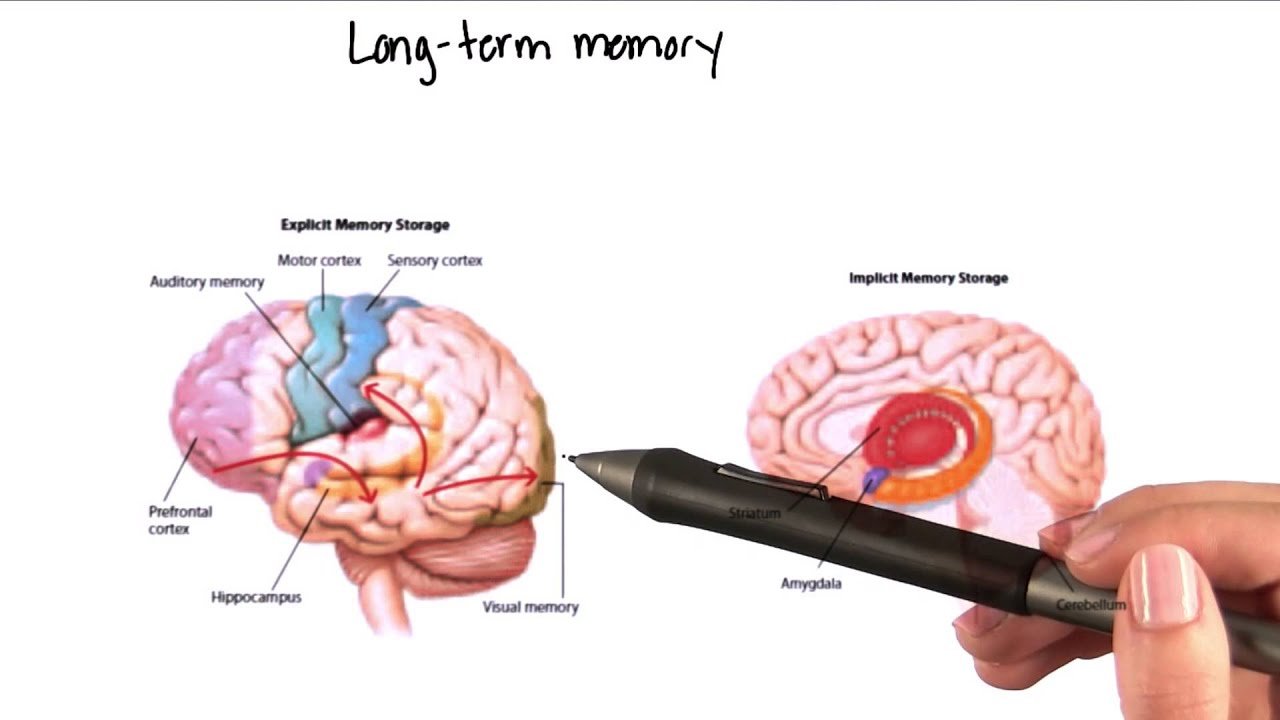
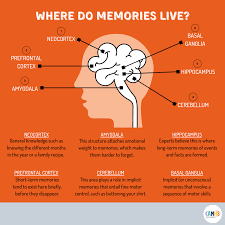
Neuroscientific research highlights the role of the hippocampal formation in coordinating this reconstruction. The hippocampus does not store the full sensory content of experiences permanently. Instead, it functions as a binding structure that temporarily links together distributed cortical representations.
When an experience occurs, the hippocampus creates associations between different neural assemblies that were active at the same time. These associations form an index that allows the brain to later reactivate the corresponding cortical networks. Without such indexing, sensory fragments would remain isolated and difficult to retrieve as coherent memories.
Damage to the hippocampus dramatically illustrates this function. Individuals with severe hippocampal injury often retain previously established memories while losing the ability to form new episodic memories. They can perceive events in the present moment, but those experiences fail to become integrated into long-term autobiographical memory.
This condition reveals that memory formation involves a transitional phase in which hippocampal coordination gradually strengthens connections among cortical regions. Over time, these cortical networks become capable of interacting directly, reducing reliance on hippocampal indexing.
The gradual redistribution of memory traces is often described as systems consolidation. During this process, patterns originally coordinated by the hippocampus become embedded more deeply within cortical networks. Sleep appears to play a crucial role in facilitating this transformation.
During certain phases of sleep, neural oscillations within the hippocampus and neocortex synchronize in precise temporal patterns. These oscillations may enable the repeated reactivation of recent experiences, allowing cortical circuits to refine the connections necessary for long-term storage.
Electrophysiological recordings reveal brief bursts of activity known as sharp-wave ripples in the hippocampus during sleep. These events coincide with coordinated activity in cortical areas, suggesting that the brain is replaying fragments of recent experiences.
Through repeated replay, neural pathways representing elements of an experience become strengthened according to principles of synaptic plasticity. Connections between neurons that activate together increase their efficiency, making future reactivation more likely.
However, memory cannot be reduced simply to strengthened synaptic connections. The structure of neural networks also influences how experiences are encoded. Each neuron participates in numerous circuits, and patterns of activity extend across populations rather than residing in single cells.
This distributed coding means that a particular memory is represented by the collective activity of many neurons. No single neuron contains the memory itself. Instead, the pattern formed by their interactions constitutes the neural representation.
The distributed nature of memory contributes to its resilience. Even if some neurons are damaged or lost, the overall pattern may still be reconstructed by the remaining network. This redundancy allows memories to survive partial disruptions in neural tissue.
At the same time, distributed representation introduces ambiguity. Because neurons participate in multiple patterns, the activation of one memory may partially activate related memories. This overlap explains why recalling one event often triggers associations with other experiences.
Emotional systems strongly influence how these patterns are formed. Structures within the limbic system evaluate the significance of experiences according to emotional relevance. Events associated with strong emotional responses often produce more robust memory traces.
The amygdala plays a central role in modulating this process. When emotionally significant stimuli appear, the amygdala influences hippocampal and cortical activity through neuromodulatory signals. These signals alter synaptic plasticity, strengthening the encoding of emotionally relevant information.
This mechanism explains why emotionally intense experiences are often remembered with unusual clarity. The neural systems responsible for evaluating emotional significance amplify the processes that stabilize memory traces.
Yet emotional intensity does not guarantee accuracy. While strong emotion enhances the vividness of recollection, it can also bias the reconstruction process. Individuals may feel certain about details that were never actually present during the original event.
Memory therefore involves a dynamic interaction between neural representation and interpretive reconstruction. Each retrieval event modifies the memory itself, subtly altering the neural pattern that will later be reactivated again.
This phenomenon is known as reconsolidation. When a memory is recalled, the neural networks representing it temporarily enter a labile state. During this period they can be modified before becoming stabilized again.
Reconsolidation suggests that memories remain biologically flexible even after long periods of stability. New information encountered during retrieval can become integrated into the memory trace, changing its structure.
The brain’s predictive architecture further complicates the question of memory location. Neural systems continuously generate predictions about sensory input based on prior experience. These predictions shape perception by influencing how incoming information is interpreted.
Memory thus contributes not only to recollection of the past but also to construction of the present. When sensory signals arrive, they are compared with stored patterns representing previous experiences. Perception emerges from the interaction between prediction and incoming data.
Because of this predictive function, memory is embedded within perceptual systems themselves. Visual cortex, auditory cortex, and other sensory areas contain traces of past experiences that influence how new stimuli are processed.
For example, repeated exposure to certain visual patterns can modify neuronal responsiveness in visual cortex. These modifications reflect learning at the level of perception, demonstrating that memory is integrated within sensory processing circuits.
Motor systems also contain forms of memory. Skills such as writing, playing an instrument, or performing athletic movements rely on procedural memory stored within motor networks and associated subcortical structures.
Unlike episodic memories, procedural memories are often difficult to describe verbally. They are expressed through coordinated patterns of movement rather than through conscious recollection. Their neural substrates involve interactions between the basal ganglia, cerebellum, and motor cortex.
These systems refine motor patterns through repeated practice. Synaptic modifications gradually optimize the efficiency of neural pathways controlling movement, allowing actions to become automatic.
The cerebellum contributes particularly to the timing and precision of learned movements. Through complex feedback loops, it adjusts motor commands based on discrepancies between intended and actual outcomes.
Procedural memory demonstrates that memory is not restricted to conscious experience. Large portions of neural learning occur outside explicit awareness, shaping behavior without generating clear recollections.
The distributed nature of memory raises deeper philosophical questions about personal identity. If memories are not located in a single place but instead arise from patterns across the nervous system, the sense of a continuous self becomes a dynamic process rather than a fixed entity.
Autobiographical memory integrates episodic experiences with semantic knowledge about one’s life. This integration occurs through interactions between hippocampal structures and networks within the medial prefrontal cortex.
These networks help organize memories into coherent narratives. By linking individual events with broader themes, they create the sense that experiences belong to the same ongoing life story.
However, this narrative construction is not static. As new experiences occur, the interpretation of past events may change. The brain reorganizes autobiographical memory to maintain coherence with evolving identity.
The plasticity underlying these processes reflects fundamental properties of neural tissue. Synapses, dendritic spines, and molecular signaling pathways continuously adapt to patterns of activity. This adaptability enables the nervous system to encode experience but also ensures that memory remains malleable.
Molecular mechanisms contribute to the stabilization of memory traces. When neurons activate repeatedly in coordinated patterns, intracellular signaling cascades trigger gene expression that supports long-term synaptic change.
Proteins produced through this gene expression modify synaptic structure, strengthening connections between neurons that participated in the original activity pattern. These molecular events allow transient electrical signals to produce lasting biological changes.
Nevertheless, even these molecular modifications occur within networks rather than isolated cells. Memory remains fundamentally relational, emerging from interactions among numerous neural elements.
The question Where memory? therefore resists a simple anatomical answer. Memory is not confined to a single structure, nor is it stored as a static record. Instead, it exists as a dynamic pattern distributed across sensory, emotional, cognitive, and motor systems.
Each time a memory is recalled, the brain reconstructs an approximation of past neural activity. This reconstruction depends on the coordinated interaction of hippocampal indexing, cortical representation, emotional modulation, and predictive processing.
Through these mechanisms the brain transforms experience into patterns that influence perception, decision-making, and identity. Memory becomes less a place within the brain than a process unfolding across time.
Understanding this process reveals that the past is not preserved intact within neural tissue. Instead, it remains alive as a set of adaptable patterns that continually interact with present perception and future expectation.
In this sense, memory is everywhere and nowhere simultaneously. It permeates the architecture of the nervous system while existing only when patterns of activity bring fragments of experience together once again.